VCR_LTER_III Long-Term Ecological Research on Landscape
Development and Ecological Processes in a Tide-Dominated
Barrier-Lagoon Complex: LTER II NSF DEB 92-11772; $1,241,406;
September 1, 1992 to October 31, 1994.
VCR LTER II focused on ecosystem change:
disturbance, succession, and ecosystem state change. We addressed the
following hypotheses: 1) The availability of fresh water, the salinity
of available water, the frequency of tidal inundation, and the
dynamics of sediment deposition control succession at the VCR; 2)
Gradients in salinity, tidal inundation, organic matter accumulation,
primary productivity, and nutrients, are functions of the land slope,
sea level, and astronomical and storm tides; and 3) Ecological state
changes are triggered by systematic long-term phenomena such as sea
level rise and sedimentation, as well as by intense disturbances such
as coastal storms. Our studies are conducted at the Virginia Coast
Reserve (VCR) on Virginia's Eastern Shore 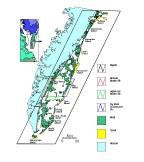 (Fig. 1). The Nature Conservancy is
the steward of our site. During the two years of LTER II, we extended
our research from LTER I, continued our systematic long-term
measurement program, initiated three long-term experiments designed to
bring about transitions in ecosystem states, incorporated new
technologies needed to obtain high resolution measurement of critical
landscape attributes (sea, land, and water table levels), and further
developed our research infrastructure in support of long-term
research.
(Fig. 1). The Nature Conservancy is
the steward of our site. During the two years of LTER II, we extended
our research from LTER I, continued our systematic long-term
measurement program, initiated three long-term experiments designed to
bring about transitions in ecosystem states, incorporated new
technologies needed to obtain high resolution measurement of critical
landscape attributes (sea, land, and water table levels), and further
developed our research infrastructure in support of long-term
research.
From January 1992 through the present, 31 journal
articles have appeared in print,14 are in press (1994 citations) and
13 are under review (1994 citations). VCR PIs have contributed 15 book
chapters in print and 4 in press and 2 books which offer synthesis of
research and theory at the site and also support intersite and network
initiatives. Twelve theses and dissertations were completed.
This
document summarizes our scientific activities for each of the 4 main
research sites at the VCR: 1) The VCR Megasite; 2) The North Hog
Chronosequence Site; 3) The Mainland Marsh Site; and 4) The Hog Island
Bay Lagoons and Marshes Site. In addition, we review the progress of
our long-term experiments; detail our accomplishments resulting from
intersite, network, modeling and synthetic studies; and identify the
relationship between past and proposed work.
Large-scale, long-term research at the VCR focuses on the causes and
effects of ecosystem disturbance and change. For example, we detailed
the hydrodynamics of the greatest coastal storm since 1942 (Davis and
Dolan 1992); we derived estimates of the wave energies of 1447 coastal
storms since 1942 (Dolan and Davis 1993) and analyzed the impact of
these storms on island dynamics (Fenster and Dolan 1994). We
calculated the occurrence probabilities of 100 barrier island plant
species across the historical storm disturbance gradient (Fahrig et al
1993)  (Fig.
5). We have also documented, using tree rings, individual tree
response to hurricanes (Johnson & Young 1992). On longer time
scales Foyle (1994), Foyle and Oertel (1994), Oertel and Kraft (1993)
and Oertel et al. (1993) charted the drainage patterns of VCR lagoons
and showed their continuity with paleochannels on the continental
shelf. These finding stimulated a new model for the origin of the
lagoon and marshes of mid-Atlantic barrier islands (Oertel and Kraft
1993).
(Fig.
5). We have also documented, using tree rings, individual tree
response to hurricanes (Johnson & Young 1992). On longer time
scales Foyle (1994), Foyle and Oertel (1994), Oertel and Kraft (1993)
and Oertel et al. (1993) charted the drainage patterns of VCR lagoons
and showed their continuity with paleochannels on the continental
shelf. These finding stimulated a new model for the origin of the
lagoon and marshes of mid-Atlantic barrier islands (Oertel and Kraft
1993). Recent efforts include increased use of remotely sensed data
as a tool for documenting ecosystem processes. We completed the GIS
framework for the annual VCR colonial bird nesting survey and have
included the 1993 GPS geo-referenced bird survey is in this system. In
the long-term we will track changes in utilization of the landscape by
colonial birds. In addition, we have mapped land cover and changes in
land cover for the VCR using TM satellite imagery. We installed a NASA
sun photometer to better correct satellite imagery for atmospheric
effects; completed our first study of spectral reflectance of barrier
island plants (Carter and Young 1993); and demonstrated the utility of
TM, Spot and aerial photography for analysis for vegetation change
(Porter and Callahan 1994).
Habitat composition and utilization at
this large spatial scale remain strong research areas. Biogeographic
surveys of island vegetation, lagoonal fish (Yozzo 1994), marcoalgae
(Monti 1993), formainifera (Woo 1992), and insular small mammal
populations (Porter and Dueser 1994, Scott and Dueser 1992, Halama and
Dueser 1992) were completed in late 1993. Tissue samples were
collected for allozyme analysis of small mammals on 11 islands;
analysis of blood samples to test for the presence of Hantavirus were
negative (Moncrief, pers. comm.). Forys and Dueser (1993) documented
movements of rice rats (Oryzomys palustris) between Crescent
and Parramore Islands, all of which originated on the smaller island.
We found that rats dispersing between islands comprise a statistically
random subset of the resident population with respect to age and sex,
unlike overland dispersers which are predominantly juveniles. Our
biogeographic surveys are conducted on a campaign basis, with expected
surveys at intervals of 3-5 years.
Harris (1992) identified and
dated a chronosequence of dune and swale landscape elements between
the ocean and the lagoon behind Hog Island. The youngest features are
located near the active beach, and the oldest (dating from ~1871) are
adjacent to the lagoon-side marsh 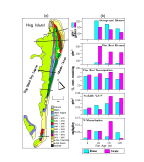 (Fig. 2A). Several studies indicate that
dune age is an important control on ecological development. For
example, soil nitrogen (ammonium and nitrate), net N-mineralization,
and total N-mineralization rates are all highest in the oldest terrain
and lowest in the newest landscapes (Young et al 1992) (Fig. 2B).
Decomposition rates are greater in older landscapes than in newer
terrain, and total below ground biomass increases from 582 g/m2 to 3035 g/m2 with increasing age of swales
through the 120 year chronosequence. Above ground and below ground
production on the dunes decreases with increasing site age (Fig. 2B);
available nitrogen increases with age in the chronosequence while
nitrogen mineralization is not well explained by landscape age (Fig. 2B). Continued chronosequence
research will focus on long-term responses of primary production,
organic turnover, and nutrient cycling rates to changes in island
hydrology, nutrient availability, herbivory and storm events.
(Fig. 2A). Several studies indicate that
dune age is an important control on ecological development. For
example, soil nitrogen (ammonium and nitrate), net N-mineralization,
and total N-mineralization rates are all highest in the oldest terrain
and lowest in the newest landscapes (Young et al 1992) (Fig. 2B).
Decomposition rates are greater in older landscapes than in newer
terrain, and total below ground biomass increases from 582 g/m2 to 3035 g/m2 with increasing age of swales
through the 120 year chronosequence. Above ground and below ground
production on the dunes decreases with increasing site age (Fig. 2B);
available nitrogen increases with age in the chronosequence while
nitrogen mineralization is not well explained by landscape age (Fig. 2B). Continued chronosequence
research will focus on long-term responses of primary production,
organic turnover, and nutrient cycling rates to changes in island
hydrology, nutrient availability, herbivory and storm events.
Through extensive surveys we have established the relationship between
perched freshwater reserves and local land elevation across scales
from microtopographic variations on marsh surfaces to the scale of
dune/swale systems. In this framework, coastal storms transport sand
and elevate land surface levels, thereby permitting higher fresh
water-table elevations (Stasavich and Hmieleski 1993). Interestingly,
over 90% of the beach sand washed inland by storm events remains in
place on the island; only 10% of the sand returns to the beach by
winds (Cohn 1993; Clark 1993). In areas of Hog Island where storm
overwash disturbance is frequent, clonal growth forms dominate (Fahrig
et al. 1993). Both ocean side flooding due to storm surge and lagoon
side flooding due to wind tides result in salinity stresses on
terrestrial island vegetation. Young and co-workers have studied the
effects of salinity on barrier island flora. Young et al. (1994)
established the salitinty tolerances of individual plant species.
Johnson & Young (1992) determined the sensitivity of pines to salt
water flooding and the effects of pore-water salinity on germination
(Sande & Young 1992) and primary productivity (Young 1992). These
studies highlight the sensitivity of the VCR ecosystem to subtle
changes in the relative elevations of the land surface and the
fresh/salt water interface.
Our studies of marsh dynamics at the landscape
level have lead us to propose a conceptual model of sea-level induced
transition from terrestrial forest to estuary (Brinson et al. 1994).
Important components of this model include self-maintaining properties
and state changes due to disturbance or exposure to acute stress. The
mechanisms responsible for maintenance and transition along the
gradient from lagoon to upland are unique for each zone within the
marsh. For example, the position of the upland-high marsh transition
depends upon the slope of the marsh surface and the distance from a
tidal creek. Along steep marsh surface slopes, elevation is the
primary factor influencing the position of the transition, while along
very gentle slopes distance from the nearest tidal creek is most
important. Along the gradient from low to high marsh at the
mainland marsh site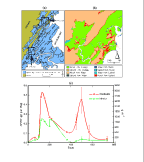 (Fig. 3A), root decay processes
apparently are not affected by sediment physico-chemical differences,
although root production is much greater in the mid-marsh zone perhaps
due to tidal inundation conditions. Greater root production and
slower root turnover occur in the mid-marsh sediments with highly
variable salinities, high oxidation-reduction potentials, lower
sediment saturation, and high sulfide concentrations. Differences in
organic matter accumulation between high and low marsh areas are thus
due to differences in root production rather than root decomposition
(Blum 1993). Although position in the marsh and sediment pore water
chemistry has little effect on root decay, Juncus roots decay
two times faster than Spartina roots. This difference is
consistent with the difference in the starting C:N ratios of 37:1 and
47:1 for Juncus and Spartina, respectively. We are
comparing two methods for measuring below ground decay: (1) decay in
undisturbed plots in which new organic matter production is prevented
by removing above ground plant material and pruning roots to exclude
new root growth, and (2) decay in buried litter bags -- and conclude
that regardless of the type of organic matter (i.e., Juncus or
Spartina) or location in the marsh, the vast majority of below
ground organic matter is old and recalcitrant (Christian et al. 1993)
and/or the removal of new below ground root material restricts
decomposition (Tirrell and Blum 1992).
(Fig. 3A), root decay processes
apparently are not affected by sediment physico-chemical differences,
although root production is much greater in the mid-marsh zone perhaps
due to tidal inundation conditions. Greater root production and
slower root turnover occur in the mid-marsh sediments with highly
variable salinities, high oxidation-reduction potentials, lower
sediment saturation, and high sulfide concentrations. Differences in
organic matter accumulation between high and low marsh areas are thus
due to differences in root production rather than root decomposition
(Blum 1993). Although position in the marsh and sediment pore water
chemistry has little effect on root decay, Juncus roots decay
two times faster than Spartina roots. This difference is
consistent with the difference in the starting C:N ratios of 37:1 and
47:1 for Juncus and Spartina, respectively. We are
comparing two methods for measuring below ground decay: (1) decay in
undisturbed plots in which new organic matter production is prevented
by removing above ground plant material and pruning roots to exclude
new root growth, and (2) decay in buried litter bags -- and conclude
that regardless of the type of organic matter (i.e., Juncus or
Spartina) or location in the marsh, the vast majority of below
ground organic matter is old and recalcitrant (Christian et al. 1993)
and/or the removal of new below ground root material restricts
decomposition (Tirrell and Blum 1992).
Field measurements and GIS
analysis of aerial photography were used to correlate landscape
evolution to sedimentary processes at the Mainland Marsh Site. The
marsh area has increased by 8% over 50 years, primarily because of
upland encroachment (Kastler 1993) (Fig.
3B). Lead dating of the sediments indicate a sediment accretion
rate of 2 mm per year (Kastler 1993), which is consistent with earlier
measures by Barr (1989) and Oertel (1992). Analysis of the clay
mineralogy of marsh sediments indicate that this marsh developed on
top of upland soils similar to those found in neighboring agricultural
and forested areas (Robinson 1993).
At the Mainland Marsh Site and
in other VCR tidal creeks, the rate of carbon cycling by bacteria is
much lower than in nearby creeks of Chesapeake Bay despite high
concentrations of inorganic N and P (MacMillin et al. 1992). The high
inorganic nutrient pools combined with the low levels of bacterial
productivity indicate that bacterial production is not limited by N or
P and that the amount of carbon moving through the bacterial loop is
low relative to Chesapeake Bay tidal creeks on the Delmarva Peninsula.
Comparative studies of VCR marshes and tidal creeks with those of
Chesapeake Bay will continue through funding from NOAA Sea Grant
program. The Phillips Creek area in the Mainland Marsh Site will
continue to be a focal point for research into the processes that
govern the hydrology, geochemistry and ecology of the marsh system.
Textural and
mineralogical analysis of inorganic sediments from throughout the VCR
revealed that each physiographic area (i.e., mainland, marsh surface,
channel bottom) has a distinct sedimentological signature (Oura 1993;
Robinson 1993). Regions of elevated paleotopography are identified as
drowned uplands through the presence of kaolinite and vermiculite
(similar to mainland soils) sampled in meter-deep cores. Preliminary
210Pb dating of sediment cores suggests that
sedimentation rates in the VCR system are 1.0-2.2 mm/yr (Kastler
1993); major storm horizons are easily identified within the shallow
cores (Kastler 1993; Robinson 1993). Sediment texture places
important constraints on the productivity of individual marsh plants.
For example, sandy creek-bank areas that are flushed by semi-diurnal
tides are highly productive; in contrast, sandy areas that are not
subject to twice daily inundation achieve hypersaline conditions and
have low Spartina alterniflora productivity (Robinson 1993). In
addition, Hussey and Odum (1992) found substantial differences in
marsh evapotranspiration (ET) across salinity gradients; the highest
ET and LAI are measured in marshes with low salinity, suggesting that
above ground primary production is salinity dependent. Extractable
ammonium and porewater ammonium and phosphate in the surface layers of
young salt marshes (10-13 years) are nearly identical to those in
marshes >100 years old, indicating a more rapid chemical maturation
than previously determined (Osgood and Zieman 1993a,b; 1994).
Three long-term
experiments were installed during 1992 and 1993. These experiments are
designed to modify the relative levels of the land, the fresh water
table and the lagoon waters. In our marsh-surface lowering experiment,
sections of the marshes were lowered 15 cm to mimic a sea level change
in excess of upward accretion of the marsh surface. Results from the
first year indicate that biomass, stem density and stem heights all
increased as a result of the relative increase in lagoon elevations
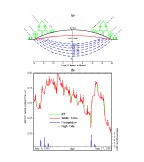 (Fig.
4A). A method of lowering the marsh surface 5 cm has been
developed and will be used in additional marsh lowering sites. The
second long-term experiment, installed in June 1993, involves a solar
powered pump to lower the local fresh-water table by 1 to 3 mm d-1. During 1993, we did not alter the water table but pumped
back into the research site and observed the natural variations in
water table at mm d-1 resolution. We have determined that
we can monitor changes in the water table resulting from semi-diurnal
tides, spring-neap tidal cycles, daily evapotranspiration and rainfall
input (Fig. 4B). Our third experiment
is a sea level manipulation. We installed (August 1993) a system of
retaining walls across a marsh ecotone and a solar powered sea water
pumping system that will permit modest changes in the duration of
tidal flooding similar to those that are occurring on longer time
scales with the current rate of sea level rise.
(Fig.
4A). A method of lowering the marsh surface 5 cm has been
developed and will be used in additional marsh lowering sites. The
second long-term experiment, installed in June 1993, involves a solar
powered pump to lower the local fresh-water table by 1 to 3 mm d-1. During 1993, we did not alter the water table but pumped
back into the research site and observed the natural variations in
water table at mm d-1 resolution. We have determined that
we can monitor changes in the water table resulting from semi-diurnal
tides, spring-neap tidal cycles, daily evapotranspiration and rainfall
input (Fig. 4B). Our third experiment
is a sea level manipulation. We installed (August 1993) a system of
retaining walls across a marsh ecotone and a solar powered sea water
pumping system that will permit modest changes in the duration of
tidal flooding similar to those that are occurring on longer time
scales with the current rate of sea level rise.
Our
theoretical, synthesis and modeling papers focus on the ecosystem
response to global climate change (O'Brien et al. 1992; Ray et al.
1992; Shugart 1992a, b, c; Shugart, Leemans, and Bonan 1992a, b, c;
Shugart and Prentice 1992, Shugart and Smith 1992b; Shugart, Smith and
Post 1992; Smith et al. 1992a, b, c; Shugart 1993a, b; Shugart and
Soloman 1993, Smith and Shugart 1993; Soloman and Shugart 1993; Davis
et al. 1994; Hayden 1994a, b, c; Shao et al. 1994a, b); succession in
terrestrial and upland ecosystems (O'Brien et al. 1992; Shugart 1992;
Smith et al. 1992a and b; Solomon and Shugart 1992; Urban et al.
1992a, b; Friend et al. 1993; Fahrig et al 1993 (Fig. 5) and
1994; Lauenroth et al. 1993; Shao et al. 1993, 1994a, b); and on the
structure and dynamics of coastal and pelagic ecosystems (Ray and
Hayden, 1992a, b; Oertel 1993a, b, 1994, Oertel et al. 1994a, b, c;
Christian 1994). Some of these efforts directly involved work for
another LTER site (O'Brien et al. 1992; Lauenroth et al. 1993) or
involved authors from another site (Shugart, Leemans, and Bonan 1992a,
b, c; Smith et al. 1992; Lauenroth et al. 1993). We have also
participated directly in providing published overviews of integrating
regional models of ecosystems with social systems (Hayden 1994a) and
of long-term data needs for long-term studies and modeling efforts
(Hayden 1994 b).
The work completed to date, within the context of prolonged scientific
discussion among the VCR PIs, has prompted the development of a
stronger theoretical foundation for our proposed research (see Section
1: Project Description, pages PD 5-6). This is the most significant
accomplishment of the last year and a half as it has brought all PIs
into a common research framework. Studies reported on in this section
will continue in the 6 years of proposed work.
Since 1992, we improved our
research infrastructure at both the UVa laboratories and at the VCR
site. We have improved our laboratories for remote sensing, GIS and
modeling with new equipment and equipment upgrades. In addition, we
added new capabilities in the area of data management and information
exchange (see Section 2, page PD 27). Stephen Macko joined the LTER
PI team and brings a significant capability in the area of organic
geochemistry and isotope biogeochemistry (see pages APDX 3 - 4). New
PI James Galloway brings to the VCR LTER his analytical laboratory for
precipitation and water chemistry and his field wet and dry deposition
station on Hog Island. The addition of Tanya Furman gains VCR access
to her mineralogical and x-ray diffraction laboratory. Added PIs
Patricia Wiberg, G. Carleton Ray, Jerry McCormick-Ray and Iris
Anderson bring expertise in sediment dynamics, marine mammal ecology,
invertebrate ecology and trace gas emissions. We also air conditioned
our site laboratory and lodging areas, built a new outdoor,
screened-in sample processing facility, purchased a mm-resolution
laser surveying system, and completed our cm (x,y,z)-resolution,
kinematic GPS surveying system.
Between 1992 and the present we have
graduated 11 students with advanced degrees (8 MS and 3 Ph.D.) who
have received part or all of their support from the VCR LTER core
grant. We currently have 13 MS students and 5 Ph.D. students
conducting research at the site. Thirteen of the published papers
reported on in the results of prior NSF support involved graduate
students. In addition, we had 4 REU students during the 1992 field
season and following semester, and 5 REU students in 1993. Three of
the four 1992 students are now in Graduate School and the fourth is
applying to Graduate School. All of the 1993 REU students have plans
to enter graduate school. We supported 4 additional undergraduate
students in 1993 and they also have plans for graduate school.