Introduction: Sediment-water material exchanges are an intergral component of the nutrient and dissolved oxygen cycles in shallow water bodies. These fluxes represent the interactions of both biological and chemical processes and are widely evaluated as indicators of the metabolism of benthic communities (Rizzo et al. 1995, Fisher et al. 1982, Kemp and Boynton, 1981,1980). Environmental conditions such as light attenuation, wind velocity, sediment loading from erosion, and anthropogenic activities may effect the magnitude and direction of benthic material fluxes into and from the water column. We proposed to relate benthic community structure to the sediment-water material fluxes of nutrients and dissolved oxygen from sediments exposed to different light conditions. The results of this study were then compared to sediment-water fluxes measured at 3 National Estuarine Research Reserve sites. A trophic state index based on the net community productivity and community respiration was assigned to each of the sites sampled after the design of Rizzo et al. (1995).
Methods: Sediment-water material exchanges and community structure were determined in cores collected from one and two meter water depths from the northern estuarine shoals of Hog Island (HI), VA on July 10th, 1994. The sampling apparatus and experimental design were those developed and described by Rizzo et al. 1992 and Lackey 1992. Cores were incubated under 5 light treaments in 4 replicates and nutrient and dissolved oxygen samples were taken at the beginning and and of the experiment. DO was measured at the sampling site and NH4, NOx, and PO4 concentrations of the overlying water column were measured at the East Carolina University Central Environmental Laboratory. Fluxes were calculated for the nutrients (umole /m2 /h) and dissolved oxygen (mg /m2 /h). From the light incubated (0 screens) and dark incubated cores, community structure was examined through sampling the microalgae (ug /Chl a /cc), bacteria and protozoa (individuals /cc), and meiofauna (individuals /9.3 cc) from the top 1 cm of the sediment. Mean data from these light and dark incubation experiments were compared to data collected n the summer of 1993 from Goodwin Islands (GI), VA, Masonboro Island (MI), NC, and Currituck Banks (CB), NC.
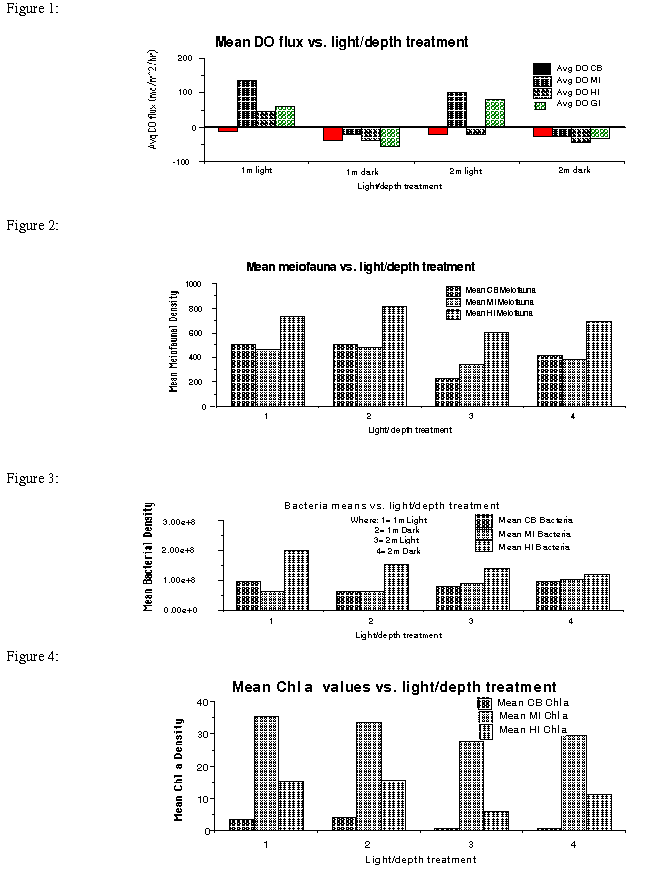
Results: Fluxes of DO2, NH4, and NOx were classified by depth and light treatment in figures representing the production and respiration of the total benthic community. In the dark incubations, DO2, was taken up by the sediments as indicated from the flux calculations (Figure 1). The magnitude of mean fluxes of all material fluxes from or into sediment of the 1-m depth were consistently greater than in cores from the 2-m water-column depth. Mean NH4 fluxes
Figure 1: DO2 fluxes by light/depth treatment where 1=1m Light, 2=1m Dark, 3=2m Light, and 4=2m Dark refers to water-column depth and light incubation treatment.
in both light and dark incubations into Hog Island sediments were higher than those measured from all of the other sampling sites except for the 2-m MI site. DO2 fluxes indicated production from light treatment cores except for the CB and two meter HI site. Community respiration, as indicated by an influx of DO2 from the water column to the sediment, was measured in all of the dark incubated cores.
Benthic community structure was described graphically for the GI, MI, and CB one and two meter sampling sites. The HI site showed consistently greater numbers of both meiofauna and bacteria (Figure 2) than all the other sampling sites. Mean microalgae measurements of HI were consistently higher across light and depth treatments than CB and lower than those found at MI (Figure 3).
The benthic trophic state index (described Rizzo et al. 1995) was a value of "3", or highly autotrophic, to all sites except CB and HI 1m sediments. These sites were assigned the "1" index which are described as net heterotrophic.
Figure 2 & 3: Shows a comparison of the mean bacterial and meiofaunaldensities for CB, MI, and HI. Where ligth/depth treatments are the same as those in Figure 1.
Figure 4: Shows mean sediment microalgae density as measured by Chl a (ug/cc). Where the light/depth treatment are the same as those in Figures 1&2.
Discussion: Sediments sampled from the northern Hog Island estuarine shoals appear to be a combination of highly autotrophic in the very shallowest parts of the estuary and net heterotrophic in the 2-m water-column depths. The 1-m water depths sampled are of high sand content and are within the euphotic zone which has allowed for the establishment of a productive microalgae sub-community. The combination of nutrient and material flux data and estimates of benthic community structure allowed for the comparison of direct comparisons of organisms with the material exchanges that may be associated with them. Meiofaunal and bacterial densities appear high and may explain the relatively high rate of uptake of oxygen in the 2-m water column depths of Hog Island. The relatively high uptake of the NH4 is probably results from activities of microalgae of the benthos which may be limited in density by this inorganic nutrient.
In this presentation, density of organisms and fluxes of materials were only compared graphically, however, further statistical analyses, which involve GLM applications in SAS, are in process with the thesis work of the author. In this manner, the variance associated with the fluxes of material can be specifically associated and controlled for with the classification variables of site, depth and site*depth (interaction). Quantification and direction of fluxes of inorganic nutrients and dissolved oxygen may regulate and be regulated by community structure. Analyses are being done to determine the effects of a the general autotrophic and heterotrophic communities on these sediment-water material fluxes.
Literature Cited:
Fisher, T. R., P.R. Carlson, and R. T. Barber. 1982. Sediment Nutrient Regeneration in Three North Carolina Estuaries. Estuarine, Coastal and Shelf Science 14: 101-116.
Kemp, Michael W. and Walter R. Boynton. 1981. External and Internal Factors Regulating Metabolic Rates of and Estuarine Benthic Community. Oceologia 51: 19-27.
Kemp, W.M. and W. R. Boynton. 1980. Influences of Biological and Physical Processes on Dissolved Oxygen Dynamics in an Estuarine System: Implications for Measurement of Community Metabolism. Estuarine and Coastal Marine Science II: 407-431.
Rizzo, W.M., S.K. Dailey, G.J. Lackey, R.R. Christian, B.E. Berry, R.L. Wetzel. 1995 (Accepted with revision). A Metabolism-Based Trophic Index for comparing the Ecological Values of Shallow Water Sediment Habitats. Estuaries.
Rizzo, William M., Gregory J. Lackey, and Robert R. Christian. 1992. Significance of euphotic, subtidal sediments to oxygen and nutrient cycling in a temperate estuary. Mar. Ecol. Prog. Ser. 86: 51-61.
Lackey, Gregory J. 1992. The effects of light and nutrient availability on primary productivity across the freshwater-saltwater interface of the Neuse River Estuary, North Carolina. Thesis-East Carolina University Biology Department.
{kind=link}