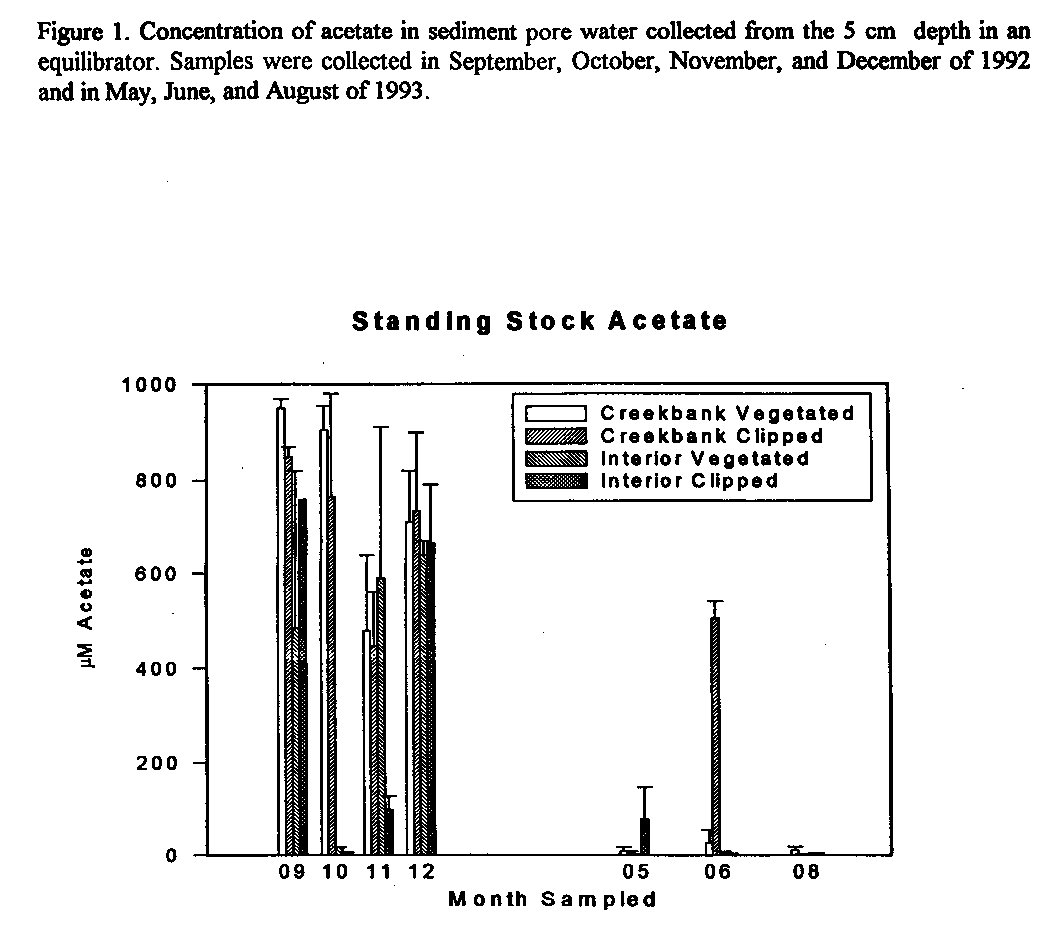
 Figure 1. Concentration of acetate in sediment pore water collected from the 5
cm depth in an equilibrator. Samples were collected in September, October,
November, and December of 1992 and in May, June, and August of 1993.
Figure 1. Concentration of acetate in sediment pore water collected from the 5
cm depth in an equilibrator. Samples were collected in September, October,
November, and December of 1992 and in May, June, and August of 1993.
Observations in Virginia, Massachusetts, and North Carolina salt marshes have shown the presence of live roots stimulate belowground decay. Hines et al. (199 ) noted increased concentrations of acetate in marsh sediments, but only when Spartina alterniflora was actively growing. They suggested that the roots were excreting acetate which was responsible for enhancing the activity of root-associated microorganisms. Acetate is an important intermediate in anaerobic decomposition both as a fermentation product and as an organic substrate for sulfate-reducing and methanogenic bacteria. If roots excrete acetate or acetate is produced in the rhizosphere via fermentation thereby stimulating overall metabolic activity of the microbial community, then it would be reasonable to expect that decomposition might be enhanced in the presence of live roots. Therefore, it was the objective of this study to determine if acetate metabolism and turnover in salt marsh sediments are enhanced in the presence of live S. alterniflora roots.
The aboveground vegetation was clipped from three plots in both the interior (short-form S. alterniflora) and creekside (intermediate-form S. alterniflora) marsh. Plastic garden edging was inserted into the marsh sediment to a depth of 15 cm to discourage root growth into the plots. Any regrowth of aboveground vegetation was removed at biweekly intervals. These clipped plots (no live roots) were established 18 months prior to the beginning of the experiments. At each location, three plots receiving no treatment were also established (live roots). Acetate mineralization was measured by injecting the sediments with tracer amounts of 14C-acetate and after an a 2-4 hour incubation, measuring the amount of 14CO2 produced. Sediment acetate in the micropore water was sampled using equilibrators. Acetate concentrations were measured using an enzyme-based high-pressure liquid chromatography assay which couples synthesis of acetyl coenzyme A to AMP production (King 1991).
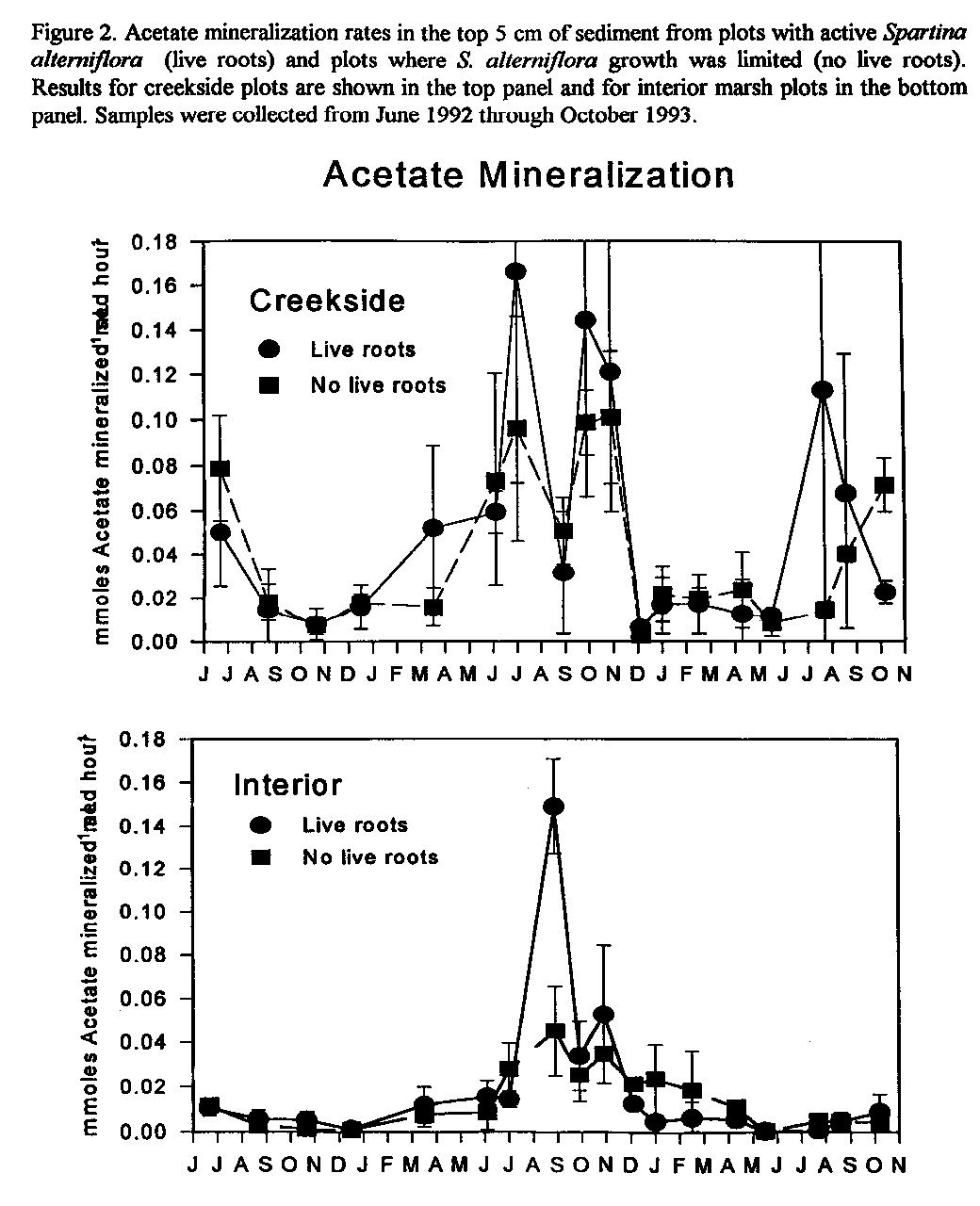
Sediment pore water acetate concentrations were greater in the winter than the summer and ranged from < 0.001 mM to > 1.0 mM. Acetate concentrations were always greater in the creekside sediments than in the marsh interior (Fig. 1). Similarly, acetate mineralization rates were greatest in creekside than in interior sediments regardless of the presence or absence of live roots (Fig. 2). Acetate turnover rates were similar at both locations and were most rapid in the summer. These data suggest that acetate mineralization by interior marsh sediment microorganisms appears to be limited by acetate supply.
In contrast to the results of others, we found that live S. alterniflora roots had no measurable impact on acetate concentrations or mineralization rates, but that flooding frequency and related physical/chemical factors may be important determinants of acetate biogeochemistry.
Figure 1. Concentration of acetate in sediment pore water collected from the 5
cm depth in an equilibrator. Samples were collected in September, October,
November, and December of 1992 and in May, June, and August of 1993.
Figure 2. Acetate mineralization rates in the top 5 cm of sediment from plots
with active Spartina alterniflora (live roots) and plots where S.
alterniflora growth was limited (no live roots). Results for creekside
plots are shown in the top panel and for interior marsh plots in the bottom
panel. Samples were collected from June 1992 through October 1993.
Click here for Picture